
Ankylosauridae - Ankylosauridae
| Anquilosaurios |
|
|---|---|

|
|
| Esqueleto montado de Scolosaurus thronus , Museo de Paleontología Royal Tyrrell | |
|
clasificación cientifica |
|
| Reino: | Animalia |
| Filo: | Chordata |
| Clade : | Dinosaurio |
| Pedido: | † Ornitischia |
| Suborden: | † Anquilosauria |
| Familia: |
† Ankylosauridae Brown , 1908 |
| Especie tipo | |
|
† Ankylosaurus magniventris Marrón, 1908
|
|
| Subgrupos | |
|
|
| Sinónimos | |
|
|
Ankylosauridae ( / ˌ æ ŋ k ɪ l oʊ s ɔː r ɪ d i / ) es una familia de dinosaurios blindados dentro Ankylosauria , y es el grupo hermano de Nodosauridae . Los anquilosaurios más antiguos que se conocen datan de hace unos 122 millones de años y se extinguieron hace 66 millones de años durante el evento de extinción del Cretácico-Paleógeno . Estos animales eran principalmente herbívoros y cuadrúpedos obligados, con dientes en forma de hoja y cuerpos robustos cubiertos de escudos. Los anquilosaurios poseen un hocico claramente abovedado y corto, osteodermos en forma de cuña en el cráneo, escudos a lo largo del torso y un garrote en la cola.
Ankylosauridae se conoce exclusivamente en el hemisferio norte, con especímenes que se encuentran en el oeste de América del Norte, Europa y el este de Asia. Los primeros descubrimientos dentro de esta familia fueron del género Ankylosaurus , por Peter Kaiser y Barnum Brown en Montana en 1906. Brown pasó a nombrar Ankylosauridae y la subfamilia Ankylosaurinae en 1908.
Anatomía
Los anquilosaurios son dinosaurios robustos, de estructura sólida y blindados. Poseen osificaciones accesorias en huesos craneales que cubren algunas aberturas del cráneo y forman estructuras en forma de cuerno en forma de cuña. A lo largo del torso de anquilosaurios hay filas de escudos, que se rellenan con huesecillos más pequeños para crear un escudo de armadura fusionado. Solo se pueden encontrar dos collares de placas de armadura en el cuello, a diferencia del grupo hermano, los nodosáuridos, que tienen tres. Nodosauridae y Ankylosauridae también comparten el atributo único de abundantes fibras estructurales en el hueso primario y secundario. Los anquilosaurios también tienen un pasaje narial en forma de S.

La característica más distintiva de los anquilosáuridos es la presencia de un garrote en la cola . Está hecho de vértebras caudales distales entrelazadas modificadas y osteodermos bulbosos agrandados. El "mango" del club de la cola involucra las vértebras y requiere que las prezigapofisis alargadas se superpongan al menos la mitad de la longitud del centro vertebral precedente. Estas vértebras caudales distales también carecen de apófisis transversales y espinas neurales y, por lo tanto, tienden a ser más largas que anchas; lo contrario es cierto para las vértebras caudales proximales. Los anquilosáuridos derivados poseen una fusión de vértebras dorsales posteriores, sacras y, a veces, caudales anteriores, que forman una estructura singular denominada "complejo sinsacro". Existe una fusión completa entre el centro, los arcos neurales, las cigapófisis y, a veces, las espinas neurales.
En 2017, Victoria M. Arbor y David C. Evans describieron un nuevo género de anquilosaurina que conservaba extensos tejidos blandos a lo largo del cuerpo. Este animal, llamado Zuul por su parecido con el monstruo Cazafantasmas , es también el primer anquilosaurio de la Formación Judith River.
Historia de estudio
Barnum Brown y Peter Kaisen descubrieron el primer género de anquilosaurios, Ankylosaurus , en 1906 en Hell Creek Beds en Montana. El material fósil que encontraron fue una porción del cráneo, dos dientes, algunas vértebras, una escápula distorsionada, costillas y más de treinta osteodermos. La reconstrucción del espécimen fue recibida inicialmente con escepticismo por aquellos que creían que era al menos muy cercano o completamente parte del género Stegopelta , y el propio Brown lo colocó dentro del suborden Stegosauria.
Se ha interpretado previamente que la variación en la forma del club de la cola del anquilosaurio se debe al dimorfismo sexual , que asume que la morfología del club de la cola tiene una función intraespecífica ligada al sexo . Esto es posible si se utilizó el palo de la cola para un comportamiento agonista. Sin embargo, una teoría del dimorfismo sexual predeciría aproximadamente el mismo número de individuos con dos tamaños distintos de garrotes de cola. El dimorfismo sexual obvio no se ha documentado, pero si los clubes de un sexo son mucho más grandes, entonces habría un sesgo de preservación y descubrimiento hacia ese sexo.
Filogenia
En 1978, WP Coombs, Jr. clasificó casi todas las especies válidas de Ankylosauria dentro de Nodosauridae o Ankylosauridae. Este fue un estudio fundamental y describió muchos caracteres de los anquilosaurios en los primeros análisis filogenéticos del grupo.
Más tarde, en 1998, Paul Sereno definió formalmente a Ankylosauridae como todos los anquilosaurios más estrechamente relacionados con Ankylosaurus que con Panaplosaurus . Los anquilosaurios que no poseían un club de cola se incluyeron en la filogenia de Ankylosauridae de Kenneth Carpenter de 2001.
En un estudio realizado en 2004 por Vickaryous et al., Gargoyleosaurus , Gastonia , y Minmi se registraron como anquilosáuridos basales, con el resto de los anquilosáuridos rellenado con gobisaurus , shamosaurus y ankylosaurines de China, Mongolia, y América del Norte.
En 2012, Thompson et al. llevó a cabo un análisis de casi todos los anquilosaurios válidos y taxones externos conocidos en ese momento. Basaron su filogenia resultante en caracteres que representan la anatomía craneal, poscraneal y osteodermal, y detalles de sinapomorfias para cada clado recuperado. Este estudio colocó a Gargoyleosaurus y Gastonia dentro de los Nodosauridae basales, y puso a Cedarpelta y Liaoningosaurus como anquilosaurios basales.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Más recientemente, Arbor y Currie han presentado un análisis filogenético de Ankylosauridae que consiste en Gastonia , Cedarpelta , Chuanqilong , otros anquilosáuridos basales y varios anquilosáuridos derivados. Su filogenia incluye algunas relaciones filogenéticas inciertas entre Ankylosaurus , Anodontosaurus , Scolosaurus y Ziapelta .
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología
Postura y locomoción
Los anquilosaurios probablemente eran animales de movimiento muy lento. En toda la anquilosauria, el peroné es más delgado que la tibia , lo que sugiere que la tibia soportó la mayor parte del peso del animal, mientras que el peroné sirvió como área de unión muscular. Los músculos de las extremidades posteriores de Euoplocephalus se han restaurado y la colocación de varios músculos que se insertan en el fémur tienen brazos de muy corto momento . Los músculos que se insertan en la tibia y el peroné tienen brazos de momento más largos. Este patrón de músculos retractores apunta a una locomoción elefantina, consistente con la postura columnar.
La restauración de las extremidades anteriores de Euoplocephalus demuestra similitudes con la musculatura de las extremidades anteriores del cocodrilo. Los músculos más desarrollados de la región pectoral tenían más una función de soporte de peso que una función de rotación. También se ha postulado que los carpos y metacarpianos se parecen a los de los tetrápodos con hábitos fosoriales (excavadores).
Varios músculos de la parte posterior de los anquilosaurios (dorsalis caudae, ilio-caudalis, coccygeo-femoralis brevis, coccygeo-femoralis longus, ilio-tibialis e ischio caudalis) se utilizaron para el movimiento de la cola y el club de la cola. Los anquilosaurios tienden a tener una orientación horizontal en lugar de una orientación vertical oblicua de las articulaciones cigapofisarias en las vértebras caudales libres de la cola. Esta disposición es más eficaz para la movilidad de lado a lado que para la movilidad vertical. La ausencia de musculatura para elevar la cola y esta orientación de las cigapófisis sugieren que la cola y su garrote se desplazaron paralelos y ligeramente por encima del suelo.
Biogeografia

Es difícil establecer el origen geográfico de Ankylosauridae en la actualidad. Existe una mezcla de anquilosáuridos basales tanto de América del Norte como de Asia, que continúa a través de análisis cladísticos aceptados. Parece que a mediados del Cretácico , los nodosáuridos asiáticos fueron reemplazados por anquilosaurios anquilosaurios. Algunos investigadores postulan que las anquilosaurios migraron a América del Norte desde Asia entre el Albiano y el Campaniano , donde se diversificaron en un clado de anquilosaurios caracterizados por hocicos arqueados y placas óseas craneales planas (caputegulae). No hay evidencia de anquilosaurios en Gondwana .
Variación
Dentro de los Ankylosauridae hay mucha variación individual e interespecífica en la expresión de la armadura. Sin embargo, el aspecto más investigado de la armadura de anquilosaurio es el garrote de cola. Se ha encontrado una variabilidad ontogenética e individual sustancial en la morfología de esta característica. Ha habido al menos 16 vértebras caudales incluidas en el mango del club de la cola de Pinacosaurus grangeri , y Euoplocephalus tiene un estimado de 9 a 11 caudales coosificados.
Las variaciones en la forma, el grosor y la longitud de la protuberancia de la cola se atribuyen a la variación individual, la taxonomía o la representación de diferentes fases de crecimiento. Sin embargo, existen dificultades con este último aspecto, ya que los clubes conocidos no se ajustan a una sola serie de crecimiento, aunque algunas diferencias deben ser ontogenéticas y alométricas .
Estilo de vida
La mayoría de los dientes anquilosaurios tenían forma de hoja, lo que implicaba una dieta principalmente herbívora. Sus dientes pueden ser lisos o estriados, o pueden diferir en las superficies labiales y linguales. Euoplocephalus tutus posee crestas y surcos en sus dientes que no tienen relación con sus cúspides marginales. Con su cuello y cabeza mirando hacia abajo, es plausible que los anquilosaurios se alimenten en un patrón de pastoreo.
Sin embargo, en algunas especies se han implicado hábitos no herbívoros. Se ha especulado que Pinacosaurus es un insectívoro de lengua larga parecido a un hormiguero , mientras que se ha propuesto que Liaoningosaurus es un piscívoro . Cualquiera de las dos sería una prueba excepcional de carnivoría entre los ornitisquios .

Hay algunas teorías predominantes sobre la función del club de cola de los anquilosaurios. El primero es el comportamiento agonístico dentro de una especie. En la mayoría de los vertebrados, incluidos los dinosaurios, este comportamiento va acompañado de estructuras de exhibición o combate. Algunos investigadores creen que este fenómeno habría sido inverosímil considerando que no existe un análogo de tetrápodo moderno que use la cola para este propósito. En cambio, estos paleontólogos proponen que los anquilosaurios hicieron uso de su cráneo ancho y plano para dar cabezazos entre individuos.
La segunda teoría de la función del palo de cola es la defensa contra los depredadores. Se ha postulado que el palo sería más eficaz contra los metatarsianos de un terópodo atacante.
Los huesos que forman la ornamentación craneal tienen costos fisiológicos, por lo que sería ineficaz producirlos simplemente para protegerlos contra la depredación. Por lo tanto, se ha planteado la teoría de que estos osteodermos en forma de cuña podrían apoyar una interpretación parcialmente seleccionada sexualmente .
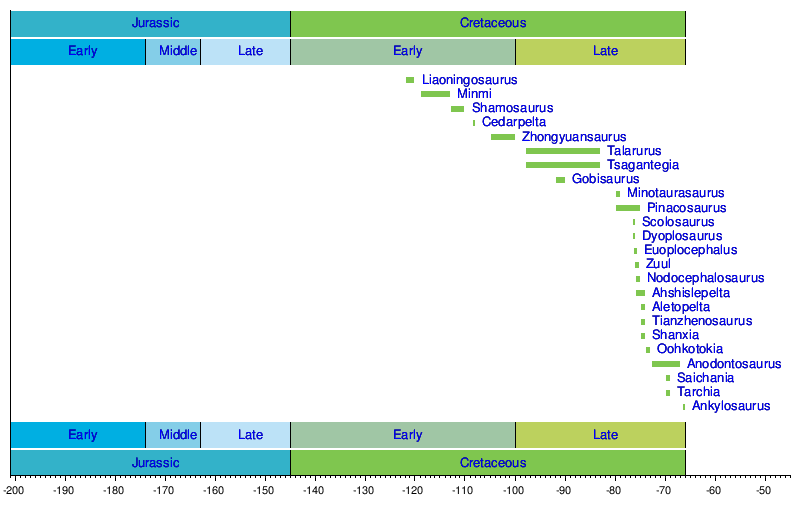
Cronología de descubrimientos

Cronología de géneros
Ver también
Referencias
- Dinosaurs and other Prehistoric Creatures , editado por Ingrid Cranfield (2000), Salamander books, pág. 250-257.
- Carpenter K (2001). "Análisis filogenético de la Ankylosauria". En Carpenter, Kenneth (ed.). Los dinosaurios blindados . Prensa de la Universidad de Indiana. págs. 455–484. ISBN 978-0-253-33964-5.
- Kirkland, JI (1996). Biogeografía de las faunas del Cretácico medio de América del Norte occidental: pérdida de los lazos europeos y el primer gran intercambio asiático-norteamericano. J. Vert. Paleontol. 16 (Supl. A 3): 45A
enlaces externos
- Árbol de familia
- Proyecto web Ankylosauridae Tree of Life
- Pseudoplocephalus , Blog de un biólogo evolutivo y paleontólogo de vertebrados especializado en anquilosaurios